花芽分化是果树连年丰产研究的重要课题,是开花结果的前提和基础,花芽分化质量的好坏、分化数量的多少与果树的开花结实密切相关;而花芽分化又是一个与植物体生长发育过程并行的发育过程,分化过程中受到的内外影响因子较多,分化时间较长。

目前,葡萄的设施栽培尤其是促早栽培已经成为鲜食葡萄早期上市的主要技术措施,具有高产出、高效益的特点,在现阶段得到大力发展。由于设施栽培环境与露地栽培环境的差异,使得一些在露地生产中发育正常的生理过程,成了制约葡萄设施栽培尤其是促早栽培顺利生长发育的关键。其中,设施葡萄不良的花芽分化状况对于其在设施产业中的发展影响最为明显,能否形成量多质优的花芽已成为葡萄设施栽培取得成功的关键。
由于人们对葡萄在设施栽培条件下的花芽分化规律缺乏足够认识,加之设施环境条件的限制和管理水平的欠缺,设施生产尤其是促早栽培中隔年结果(大小年)现象时有发生,缩短树体经济寿命,降低设施葡萄产量,增加种植者的农事操作成本,加大了投入产出比。而此种现象产生的根本原因是人们对于设施促早栽培条件下葡萄花芽分化的规律缺乏了解,对影响成花的因子缺少认识,故而明确葡萄设施栽培尤其是促早栽培花芽分化规律及成花的主要影响因子,是实现葡萄设施栽培丰产稳产的必经之路。
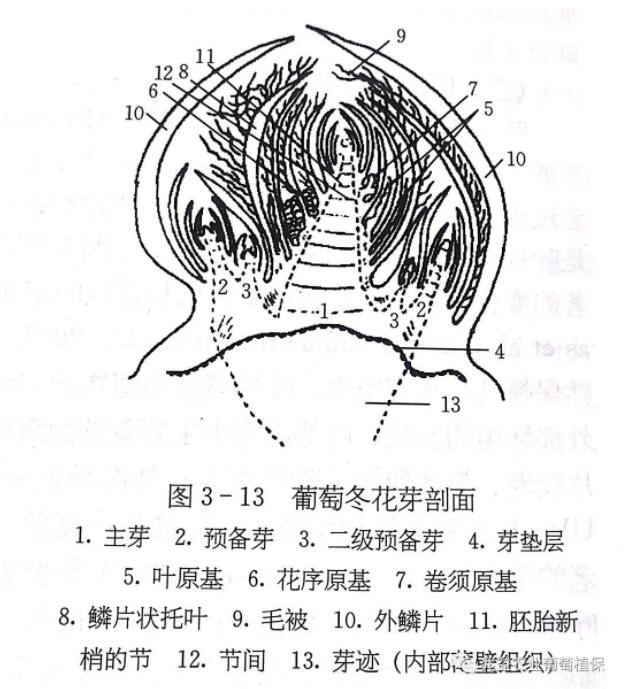
01芽的类型和形态结构 Types and morphological structures of buds 葡萄枝梢上的芽实际上是新梢的茎、叶和花的过渡性器官,着生于叶腋中,根据萌发的时间和结构特点,分为冬芽和夏芽。 (1)冬芽 冬芽是着生在结果母枝各节上的芽,体形比夏芽大,外被鳞片,鳞片上着生茸毛,保护芽体免受冬季低温的伤害。冬芽具有晚熟性,一般经过越冬后,次年春季萌发生长,但在强烈刺激下,如去除所有副梢或配合石灰氮、破眠剂等具有破眠作用的化学试剂处理,冬芽可当年萌发,在生产中常采用这种方法迫使冬芽萌发实现二次结果。 从冬芽解剖结构看,良好的冬芽一般由3~8个单芽组成,其中芽眼中央发育最好最大的单芽称为主芽,其余周围的单芽称为副芽,主芽与副芽均是压缩的新梢原基,其上有节、节间、叶原基、芽原基、花原基或卷须原基,依据发育程度,主芽的新梢原基在冬芽萌发前,可分化10~13节。 (2)夏芽 夏芽着生在新梢叶腋内冬芽的旁边,无鳞片保护,不能越冬。夏芽具有早熟性,在当年夏季自然萌发成新梢(通称副梢),有些品种如巨峰、玫瑰香、魏可和美人指等的夏芽副梢结实力较强,因此在生产中常利用夏芽的早熟性加快葡萄的成形或进行多次结果以延长葡萄鲜果的供应期。 02花芽分化 flower bud differentiation 葡萄的花芽分化可分为生理分化和形态分化两个阶段。芽的生长点分裂4~5个叶原基时,生长点转位即进入形态分化期。 决定花芽良好分化的前提首先是营养状况和外界条件(光照、温度、水份)的充分满足。花芽形成的最适温度为20~30℃,而光照充足、新梢生长健壮、叶面积大、叶片质量好,葡萄花芽分化的强度和质量也高。新梢第1~3节的芽是新梢开始生长时形成的,这时正值早春季节,气温不高,新梢生长缓慢,芽体秕小呈三角形,节间短,花芽分化不是很理想。当新梢进入第一生长高峰时,平均气温在20℃以上,是幼芽的形成和花芽分化的良好条件,这时新梢第4~5节位的芽发育最好,花芽分化率也高。由于葡萄的花芽分化与萌芽、新梢生长、开花坐果、浆果发育交叉重叠进行,因此从萌芽至开花前后及浆果膨大期需要供应充足的营养物质,同时也要进行夏季修剪(抹芽、疏枝、摘心、疏花、疏果及处理副梢),通过开源节流的措施来促进花芽分化。 如营养条件不足,有的花芽甚至退化为卷须,有的则产生不完整的花穗原基,开花后造成落花落果或无核小粒果,或产生卷须与花穗的中间产物;当营养充分时,卷须可能转化为花序,开花后果穗及果粒能正常发育。 葡萄的花芽有冬花芽和夏花芽之分,一般一年分化一次,也可一年分化多次。 (1)冬花芽的分化 葡萄冬花芽分化和发育的时间比较长,主梢开花始期也是冬芽分化始期。靠近主梢下部的冬芽最先开始分化,随着新梢的生长,新梢上各节冬芽从下而上逐渐开始分化,但最基部1~3节上的冬芽分化稍迟或分化不完全,这可能与内在生理特性和外界环境条件有关。冬季休眠期间,芽内的花穗原始体在形态上不再出现明显的变化,到第二年萌芽和展叶后,在上一年已形成的花穗原始体的基础上又继续进行分化。随着新梢生长,花序上每朵花依次分化成花萼、花冠、雄蕊、雌蕊等各个部分。 因此,树体贮藏养分的多少对早春花芽的继续分化至关重要。 葡萄生长过程中要创造一些良好的栽培措施,如主梢摘心、控制夏芽副梢生长等促进冬芽分化的过程,使在短期内形成花穗原基。故生产上也可利用逼主梢冬芽或副梢冬芽当年萌芽开花,实现二次或三次结果。 (2)夏花芽的分化 夏芽具有早熟性,一般在展叶后20d内即可成熟并萌发副梢。葡萄在自然生长状态下,夏芽萌发的副梢一般不形成花穗结果,如对主梢进行摘心,则能促进夏花芽的分化。夏花芽花穗发育的大小与夏芽萌发前的孕育时间长短有关,一般情况下,由于夏花芽分化形成的时间短,故副穗花穗较小。夏花芽的分化、结实力还因品种而异,巨峰一般约有15%的夏芽副梢有花穗。由于在年生长周期内,夏芽副梢可以多次萌发,因此可以多次开花结果,形成二次果、三次果等。葡萄生产中经常有应用这一原理进行二次果生产,以达到增产增值的目标。 设施栽培条件下葡萄的花芽分化规律与露地栽培显著不同。在设施促早栽培条件下,由于设施促早栽培条件改变了葡萄固有的生长规律,设施内梢形成的冬芽和夏芽由于受设施内环境条件特别是光照和营养的影响,花芽分化不良,不能形成足够数量的高质量花芽,因而导致葡萄果实产量逐年降低,出现“退化”现象,严重影响了葡萄的连年丰产。 (三)芽的异质性 葡萄芽的异质性是指由于品种、枝蔓强弱、芽在枝蔓上所处的位置和芽分化早晚等的不同,造成结果母枝上各节位不同芽之间质量的差异。一般主梢枝条基部1~2节的芽质量差,中、上部芽的质量好,例如生长势较旺的巨峰和夏黑等品种,中部5~10节的芽眼发育完全,大多为优质的花芽,下部或上部的芽眼质量较次;距中部向上或向下的芽眼,愈远则质量愈差。如果栽培措施得当,巨峰和夏黑等品种枝条基部1~2节也能形成良好的花芽。 03设施葡萄花芽分化进程 Facility grape flower bud differentiation process 设施葡萄从未分化到花穗原始体的花芽分化进程分为6个阶段:生长点未分化期、生长点半球/平顶期、生长点顶分期、带有苞片始原始体出现期、花序主轴及小穗原基发育期、花序二级轴(小穗穗轴)发育期(图7-1)。 花序形成起始的判定 露地栽培葡萄以始原始体出现(生长点一侧产生突起)作为花序形成起始的判定标准。此判定标准在设施葡萄促早栽培中不具有适用性,这是因为花序和卷须是同源器官,在露地栽培条件下,葡萄冬芽的始原始体比较容易向成花方向发育,而在设施促早栽培条件下,不耐弱光的葡萄品种夏黑冬芽的始原始体更倾向于形成卷须,因此,在设施促早栽培条件下,判断葡萄冬芽成花与否的标准应该以其花序主轴的出现为准。 花芽分化的关键时期 葡萄冬芽雏梢发育到含有两个叶原基时开始进入诱导葡萄成花的关键时期一生理分化期,直至始原始体出现,形态分化期即结束。赵君全(2014)研究发现,设施促早栽培的不耐弱光的葡萄品种夏黑和耐弱光的葡萄品种京蜜在初花期开始进入旺盛的始原始体分化阶段,因此初花前是诱导设施葡萄成花的关键时期-生理分化期。 葡萄冬芽内雏梢连续分化出6个叶原基后,就不再具有成花能力(Buttrose,1969)。中国农业科学院果树研究所研究发现,设施促早栽培夏黑和京蜜2~3节位多数冬芽分化出6个叶原基的时间在果实软化/转色之前的果实膨大期,因此果实软化/转色之前的内外因素密切影响着设施葡萄2~3节位冬芽的成花发育。 不耐弱光的夏黑,冬芽旺盛的营养生长是限制其在设施促早栽培条件下成花进而造成“隔年结果”现象发生的重要原因,通过采取抑制设施葡萄冬芽营养旺盛生长的技术措施均能促进其成花,进而解决设施葡萄促早栽培中存在的“隔年结果”问题。 04影响果树成花的外部因素 Facility grape flower bud differentiation process 温度 在一二年生植物中,温度的变化具有调控成花的重要作用。而在多年生的果树中,低温并非是其成花的必须;相反的,多数北方果树的花芽分化过程都是在较高温度的5~7月进行,可见在多年生植株体内成花起始的诱导过程不需要低温作用。然而,并非果树的成花过程不受到温度的影响,研究发现生殖生长需要比营养生长更高的温度(Luis A etal.,2005),而花原基的分化过程只有在较高温度下才能进行,且利于分化的温度范围是一个阈值,在该范围内的高温对于花芽分化的进程有促进作用(Zhu et al.,1997)。温度过高,则不利于花芽分化。葡萄栽培中,可以利用夏芽或是冬芽生产二次果,可见冬季低温对于成花不是必需的;而早期的研究发现在葡萄花序形成过程中,一定时间段的高温对葡萄花序的正常形成和分化是必须的。 光照 果树多是喜光植物,在自然光照条件下能正常成花,满足人们对于果实的需求,而对于设施弱光环境适应性一般较差,尤其在促早栽培条件下,冬春季的弱光低温对成花有着极为不利的影响。前人通过对一些指标的研究找到了具有较强弱光成花能力的材料,但对于这些材料的弱光下成花机理研究了解甚少。从大量的文献中发现弱光降低光合作用,改变叶片枝干的外部形态、内部结构及一些重要的激素比例,但对这些改变对弱光逆境下花芽分化的影响也是知之甚少。赵君全(2014)研究发现,气温不是限制设施夏黑葡萄冬芽成花的真正环境因子,光照条件恶化(日照时间缩短、光照度降低、光质变劣主要是紫外线比率下降)是设施栽培非耐弱光葡萄品种夏黑冬芽不能形成高比率良好花芽的根本原因,因此光照环境条件的改善是解决非耐弱光葡萄品种夏黑设施促早栽培隔年结果现象的根本措施,主要包括延长光照时数、增强光照度、增加紫外辐射和夏季更新修剪促生冬芽新梢避开不良光照环境条件等措施。 (1)光质影响植株成花 由于设施覆盖物薄膜或是玻璃的影响,使得设施内的光质成分发生变化,进而影响到植株长势,引起营养生长与生殖生长失衡。弱光易引起幼苗的徒长。(郗荣庭,1997),在已有的研究中同样发现红光能够调控植物体内GA含量而影响到植株高度和节间长度,红外线具有反作用(李书民,2000),可见光质可以通过光受体传导和控制激素平衡来调节植物长势,进而影响枝梢营养和生殖平衡。设施葡萄栽培中,由于棚膜对紫外线的过滤作用,导致设施内紫外线含量不足,而紫外线的显著促进葡萄花芽分化的作用被减弱(王海波等,2010),因此在葡萄设施栽培中紫外灯的应用对于成花应该有促进作用。 (2)光照强度影响植株成花 研究发现光照强度不仅影响到设施中植物体的光合特性及光合产量,还可以影响光合产物的种类与分配,从而嵌入到花芽形成的多因子途径中调控成花。 葡萄栽培中,环剥提高成花率,摘心去副梢同样增加花芽分化率,可见对于成花期的营养生长的抑制,人为的改变养分流向对成花发育有重要的促进作用。设施促早葡萄花芽分化的盛期正是设施内养分竞争关键时期,是储藏养分与光合养分的转换期,此时叶片的有限光合产物分配对于花芽的形成具有重要的意义。另外,有研究发现,弱光还会影响到根的活性,进而影响激素平衡。 (3)光照时间影响植株成花 各个品种的葡萄,在不同日照长度条件下,花芽形成的量也有差异,已有研究发现,日照时数显著地影响到葡萄的成花过程,尤其在其始原始体形成之后,较长的光照时间加速花序原始体的形成,而短日照延长二分枝原始体持续的时间,进而延长了其花序发育的时间,不利于花序原始体的形成。 对于长日照植物,光照时间的延长可以提前成花,浦正明等(2004)发现光照时间的延长促进草莓的花芽分化。设施促早栽培过程中,无可避免地要加覆盖措施以增强设施的保温性能,此操作会缩短设施内的光照时间,进而影响设施果树的成花过程。 矿质元素 矿质元素在果树成花的过程中,具有保持细胞液的正常状态,维持膨压平衡和能量供给、信号的传递等作用。氮肥在生产中具有双重作用,适量的铵态氮能够提高花芽分化的量,而硝酸盐则是一个有利于碳生成氨基酸的信号分子。氮肥不足则蛋白酶和结构蛋白等合成不足,影响植物各项代谢反应,直接影响花芽分化;过量则使营养生长与生殖生长失衡,亦难成花。花芽分化过程也是一个耗磷过程(赵文东等,2006),研究发现花芽分化很大程度上被有效磷水平左右(马焕普,1987),故在生产中应充分供应。钾和钙在分化过程中也具有重要的调节作用,Manochai等(1999,2005)。钙除了作为结构成分之外,还是第二信使,与钙调素一起调节花芽的分化,且能诱导GA的表达,促进花的发育。彭抒昂等(1998)在梨成花过程中钙与核酸的动态研究中发现花芽分化之前,有一个钙积累的过程,而在成花发育期,只有生长点的分生组织消耗钙并从附近的组织夺取钙。孔海燕等(2003)离体培养外施钙浓度变化对花芽分化的早期诱导有巨大影响,并认为内源钙水平是光周期对成花诱导重要的制约因素。 05关于果树花芽分化的学说 Theory on the flower bud differentiation of fruit trees 一、C/N学说 20世纪初,前人通过对大量试验分析提出了成花的C/N学说,认为C/N值高,有利于成花;反之,不利于花芽分化。该学说在一段时间之内得到了广泛的试验证实;然而,随着研究的逐渐深入,在试验中出现了一些该学说不能解释甚至完全相反的结论,人们逐渐认识到该学说存在的局限性;而一般的研究对于碳氮的具体分类比较模糊,没有明确碳素、氮素的种类及它们在成花发育过程中有重要作用,因此人们展开了一系列特定碳氮比例与成花关系的研究,结论认为:充足的糖、淀粉等碳水化合物积累对于顺利成花是有益的,这可能是因为成花过程是一个比营养生长更为耗能的过程,较高的碳水化合物不仅为成花提供结构物质也是其代谢过程中推动反应进行的能量来源。 赵君全(2014)研究发现,始原始体形成期和花序主轴形成期叶片中可溶性糖含量和淀粉含量的剧烈下降影响设施葡萄花序的形成。贮藏营养在新梢基部节位(2~3节)的冬芽成花过程中发挥重要作用,其与光合营养的良好衔接是确保葡萄花芽分化顺利进行的重要前提。 C/N学说虽然存在不足,但是其在成花生理过程研究中的作用和意义仍是不容忽视的:良好的碳水化合物供给对花芽分化的顺利进行有重要的促进作用;碳氮比值随分化进程的变化所呈现出的规律性对于植株在整个成花过程中的发育也有重要意义。因此,认为碳氮比对于成花诱导可能并非是直接作用,只是一个协助作用,其比值在一定范围内,植物体才能感受到成花刺激信号进而诱导成花。 二、激素平衡学说 激素与成花之间的关系已有很多的研究报道,从普遍的单激素作用到多种激素之间的平衡互作,众多的研究结果表明激素对花芽形成发育至开花结实的整个过程都有着重要调节作用。通过对试验结果的汇总分析,发现细胞分裂素在多数情况下会促进果树的成花过程,而GAs则是普遍的果树成花抑制激素(GA4作用除外)。这明显不同于一二年生植物中GAs促进成花的过程,可见GAs在不同类型的植物体中,其具体作用是有差别的,因而,对于一二年生植物或是模式植物的研究中发现的一些激素调节成花规律不一定适用于果树。随着研究的逐渐深入,IAA(吲哚乙酸)和ABA(脱落酸)对花芽分化作用的研究也渐受重视,在某些物种或组织中,IAA是GA合成的上游调控成分,这些调控途径虽多在一二年生植物中发现,但多年生植物可能也同样发挥作用,或至少保留着IAA与GA互作关系。也有人认为后两者作用是因为调集养分或拮抗GA对淀粉的水解作用而利于成花的。Ramirez等(2004)也提出ABA诱导花芽分化的作用可能是拮抗了GA,延缓了茎的伸长来实现的。 葡萄与其他果树类似,也是在上一生长季形成花芽,在下一生长季开花结果,然而葡萄又与其他果树之间存在差别,主要是其形成的花序原基是一个可变的同源器官,极易受到环境条件的影响,在花序未形成之前都具有不确定性,所以,在其成花发育过程中存在多个关键调控时期,而各个时期又都受到激素的调控,激素种类含量的改变或者激素之间平衡关系的打破对于花序形成有着极大的影响。 三、成花素假说 菊花叶片处于短日照条件下就能引发花芽分化,苍耳通过嫁接传递某种信号而使未被长日照处理的植株开花等一系列生理现象引起了人们思考,继而提出“成花素”假说,在成花物质学说提出数十年内,人们一直未找到该种物质,直到近几年,分子生物学家通过对模式物种(如拟南芥)的研究,才大致确定它(Corbesier L.,2007),FT蛋白是人们认为最有可能的成花素成分(Jan AD et al.,2008),虽然多是在一、二年生的植物中发现FT促进成花,但人们相信在多年生植物中同样存在这一机制(Brunner et al.,2004),因为多年生植物中确实存在着FT的同源基因。同时研究发现FT蛋白也是春化作用的产物,且FT与GA在成花过程中独立起作用,在实验中发现FT蛋白的运输是在维管束中进行的,而芽中的内源生长素对于芽和茎干维管束的贯通有明显控制作用,一些研究中发现的生长素的促花效应是否与其控制维管束的形成,提高了成花素与营养物质运输通道有关。成花素是直接调节成花基因来实现花芽分化的调控,还是通过与激素或同化物相协同组成成花的多调控系统,尚未可知。 部分内容摘自《中国设施葡萄栽培理论与实践》